Written by Alejandra Feliciano

This paper takes a systematic look at the scientific literature published in relation to Washington’s South Sound prairie plant communities, specifically to examine changes in plant diversity and community composition. Has there been plant diversity loss? And what are the community assembly dynamics behind these changes?
SUMMARY: Scientific papers published to date on the phylogenetic composition of the South Sound prairies were collected to gather information on habitat loss and plant community composition. These prairies have undergone a 51% reduction in overall plant species richness in the last 130 years. Invasive species today contribute roughly 44% of total species richness. Although most species within the community remain native, only 27% of historical native species remain in the community. In terms of functional diversity, we can observe a reduction (-6.3%) in mean competitive investment, and stagnation (-0.4%) in mean stress tolerance. The largest functional change comes from an increase (+7.7%) in mean ruderality. Higher comparative stress-tolerance allotment between historical and contemporary natives suggests that, as community assembly filters shifted, those natives better adapted to the environmental abiotic regimes than exotics were selected to persist. Invasive exotics could be displaced through community interactions via two functional strategies: 1) lowering the ruderality requirement from the current dispersal filter and/or 2) introducing natives of superior ruderal fitness to those of exotic residents, as measured through leaf traits.
Range loss in South Sound Prairies: Urgency for addressing future local extinctions

Washington’s native prairie communities have undergone severe habitat loss over the last 130 years, being displaced by agricultural and increasingly urbanized land. Extant prairies persist today in small patches in a state of extreme isolation from one another (Apostol et al., 2006), disrupting metapopulation dynamics and increasing their vulnerability.
Displacement is due in part to the influence of topography on the region’s historical development. Mountainous terrain is difficult to farm and develop for residential and commercial uses, so the vast Puget Trough prairies were tilled over to make way for the region’s urban development, which continues today. Washington’s population, 75% of which is concentrated in the Puget Trough (Floberg et al., 2004), grew a whopping 13% (an additional 1.5 million people) in the last 10 years (US Census Bureau, 2020).
At an estimated 73,000 ha during the late 1800s, the Southern Puget Sound prairies were the largest and most vast western prairie-oak ecosystems north of the Willamette Valley. Of those, only 9% remained at the last survey count 19 years ago (Chappell et al., 2001). Today there are many bird (Altman, 2011; Rogers et al., 1997; Slater and Altman, 2011; Stinson, 2016), small mammal (Fimbel and Freed, 2008; Ryan, 1997; Stinson, 2005), and insect (Fazzino et al., 2011; Haan et al., 2018) species associated with this ecosystem that have become threatened due to this dramatic loss in habitat.
Concurrent with this loss in range, prairie-oak communities have undergone severe exotic plant invasions (Crawford and Hall, 1997; Dorner, 1999). Through this systematic literature review, I will explore what is known on the current taxonomic and functional diversity status of the prairies. Has plant diversity increased or decreased over time with exotic plant invasions? How has the functional composition of these plant communities changed over time? In answering these questions, I hope to gain a deeper understanding for how community assembly filters are changing in South Sound prairies, and perhaps how they might be gained for conservation efforts.
Methods

Using Web of Science (“Web of Science,” 2020), I performed searches for “South Sound Prairies” , “Southern Puget Trough Prairie”, and “Pacific Northwest endangered plants”. The first search yielded 23 results, the second yielded 15 results, the third 24 results. Of these, 8, 4, and 3 discussed topics related to South Sound Prairies, respectively. None contained plant survey data. Remaining sources were found through cross-references in these 15 relevant publications. Most of these external sources on South Sound Prairies are non-peer-reviewed reports from the U.S. Department of Natural Resources or from The Nature Conservancy, a global environmental nonprofit.
To characterize known changes in South Sound prairie flora, I have listed literature sources into three categories. “Plant surveys” refer to studies that conducted field survey work to assess community composition. “Reviews” refer to studies that conducted literature reviews and collected community composition data from published and unpublished sources. “Historical changes” refer to studies that conducted literature reviews and collected historical community composition data from published and unpublished sources (Table III‑2).

Some of the sources disagree on whether plants are absent from the community. For example, Agoseris heterophylla is listed as “absent” in Dunwiddie et al. (2006) and later “present” in a more recent publication (Dunwiddie et al., 2014). I used the species listed by Dunwiddie in the most recently peer-reviewed flora (Dunwiddie et al., 2006) and cross-referenced them with those in the Field Portable List provided in Pierce’s supplementary documentation (Pierce et al., 2017) to create a list of South Sound Prairie species which could be analyzed by functional guild (II.D.5.a), endemism, and occurrence in time. Where functional guild data by species were missing, averages by genus were used. Still, functional guild data were incomplete, so a subset of the original dataset was sampled for functional analysis (Table III‑3).

For species not recorded in the 15 prairies by Dunwiddie, the USDA’s PLANTS database (“PLANTS Database | USDA PLANTS,” 2020) was used to determine endemism. The final list was analyzed using R version 4.0.0 (“R: The R Project for Statistical Computing,” 2020). There is no relative abundance or percent cover data in the peer-reviewed literature. Cover data from Dorner (1999) was not used due to source type.
Results
Phylogenetic diversity characterization

South Sound prairies have undergone a 51% reduction in overall plant species richness in the last 130 years (Table III‑4). Invasive species today contribute roughly 44% of total species richness. Although most species within the community remain native, only 27% of historical native species remain in the community.
Functional diversity characterization & community assembly dynamics
To understand how community assembly filters may have changed over time, I analyzed the changes in the prairie’s proportional metabolic investment into competitive, stress-tolerance, and dispersal traits. From Figure III‑1, we can observe a reduction (-6.3%) in mean competitive investment, and stagnation (-0.4%) in mean stress tolerance. The largest functional change comes from an increase (+7.7%) in mean ruderality. Ruderal traits are associated with post-disturbance survival in seed form. Increased investments into seed production, increased phenotypic plasticity, and divestments in size and stress tolerance are common (Diaz et al., 2016; Pierce et al., 2013; Wright et al., 2004).






Figure III‑1: Past and present proportional investments. Inverstments in competitive (C%), stress tolerance (S%), and dispersal (R%) adaptations. Histograms plot the distribution of proportial investment of biomass production into functional traits per species throughout the community. Mean community investments are highlighted red.
Unpacking these functional shifts by endemism (Figure III‑2) we find that native functional composition in the past and present are more distant. Extant natives have a much higher proportional investment in stress tolerance than historical natives (+19.3%). To gain this advantage, it appears that a higher proportion of ruderal ability (-11.5%) than competitive ability (-6.8%) was sacrificed.
This analysis by endemism also allows us to paint a picture of an invasion niche which is functionally characterized by exotics with high ruderality in comparison with competitive and stress tolerance traits. Exotic competitive ability is near equal to native competitive ability (+0.8%). Stress tolerance is much lower for exotics than natives (-29.0%), but this is compensated by the exotic’s superior ruderal ability (+28.3%).



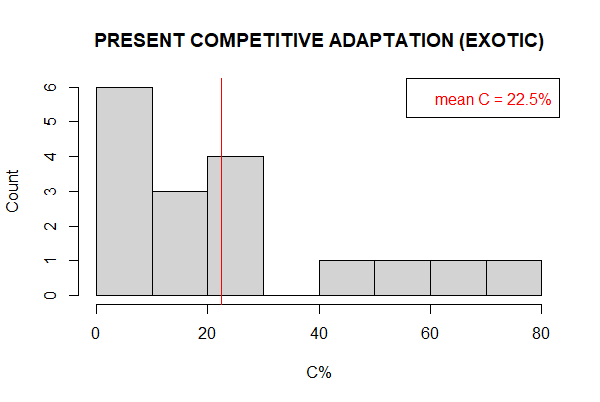


Figure III‑2: Present proportional investments by endemism. Investments in competitive (C%), stress tolerance (S%), and dispersal (R%) adaptations by endemism. Histograms plot the distribution of proportial investment in biomass production for functional traits by exotic and native species, respectively. Mean proportional investments are highlighted red.
Discussion
The state of current literature

An incomplete picture of diversity loss is presented by the current literature. Although species richness can be ascertained from field surveys, a lack of relative abundance data makes evenness assessment impossible, leading to an obscured picture of community dynamics. For example, community weighted means (Li and Shipley, 2017) of proportional functional allotment give more accurate insight into community composition and change than unweighted means, such as presented in Figure III‑1 and Figure III‑2.
An incomplete accounting of present species composition is reflected by the current literature. Many woody species reported to have encroached into prairie habitat (Peter and Harrington, 2014), such as Pseudotsuga menziesii and Rubus armeniacus, are absent from Dunwiddie’s flora (Dunwiddie et al., 2006). Such omissions cast doubts on the survey’s accuracy. In addition to these omissions, data are simply dated. The most recent survey was completed over 15 years ago, which was not peer-reviewed. Only one peer-reviewed plant survey has been performed, and that dates back over 23 years.
Of all publications related to South Sound Prairie communities reviewed in this paper, the vast majority (61%) focus on restoration strategies (Table III‑5) rather than community characterization and survey work (Table III‑2). Given the stated degradation of this ecosystem, it is understandable that the scientific community should focus on restoration strategies and plans of action. However, it is important to note that these strategies may not be successful if they are built on incomplete, dated community characterization. To develop successful restoration strategies, recent field surveys are necessary.
Shifts in community assembly filters over time
After obtaining relevant abundance data, prioritizing analysis of interactions among plants within the community and at its periphery should be a priority. Transferring rare plants and introducing them to an extirpated landscape has been shown to leave species vulnerable to competitive displacement (Clements, 2013; Dunwiddie and Martin, 2016; Ren et al., 2014).
Metabolic investments in functional traits are adaptions, meaning they occur as direct reactions to community assembly filter requirements. As such, they can provide some insight into the changing growing conditions that plant communities are exposed to over time, specifically changes in disturbance (biomass destruction), stress (biomass productivity rate), and competition (biotic interactions).
In this case, what do mean functional allotments tell us about changes in community filters over time? In comparing both past and present communities overall (Figure III‑1), the observed reduction in mean competitive ability suggests a slight lowering in community density and/or a slight reduction in the overall sizes of plants, and an increase in the frequency and/or severity of disturbance events. Unfortunately, due to lack of historical cover data, this process cannot be fully verified.
The observed stagnation in overall community stress tolerance suggests that abiotic restrictions on biomass production did not change significantly over time. Sources of abiotic stress for these prairies include gravelly glacial outwash soils with poor mineral retention, summer drought, and vernal hydric soils (Crawford and Hall, 1997; Dunwiddie and Bakker, 2011). This suggests an upper boundary in the capacity of extant prairie soils to accumulate organic matter and build more complex soils over the observed time frame.
Increased ruderality and presence of short-lived species suggests increases in disturbance over time. This seems to track with known historical disturbances (tilling, grubbing) which occurred as part of the range reduction process of these prairies.
Together these results suggest that prairie community assembly filter requirements shifted over time. They started from a more balanced, near generalist functional requirement and became skewed, selecting for more ruderal and stress-tolerant species.
Breaking down the present community by endemism provides additional insights (Figure III‑2). Higher comparative stress-tolerance allotment between historical and contemporary natives suggest that, as community assembly filters shifted, those natives better adapted to the environmental abiotic regimes than exotics where selected to persist.
Similar competitive investment between exotics and natives suggests a mechanism for coexistence, since neither appears to have competitive advantage in this regard. Superior ruderal ability in exotics seems to be associated with invasibility and colonization. Extant natives had already colonized the landscape and had less need for increased ruderality. In light of the site’s present functional state, which currently skews towards increased stress tolerance and ruderality, a mechanism for coexistence between exotics and natives could have emerged where there is a complementary trade-off between extant native’s superior stress tolerance and the invading exotic’s ruderal ability.
This hypothesis seems to track with observations of displaced annual species in the prairie (Dunwiddie et al., 2014). Almost half (42%) of historic native prairie flora is believed to have been comprised by native annuals, which are associated with high ruderal functional investments. This appears to overlap with the niche invaded by exotics, which skews towards ruderality.
Conservation Strategies
Based on the above hypothesis, any future prairie conservation plan must pay close attention to interactions involving the ruderal niche. To displace invasive exotics through community interactions, two functional strategies could be envisioned: 1) lowering the ruderality requirement from the current dispersal filter and/or 2) introducing natives of superior ruderal fitness to that of exotics.
Strategy 1: Lowering ruderality requirements
For the first strategy, manipulating dispersal filter requirements involves minimizing sources of disturbance (biomass destruction). Traditional sources of disturbance are herbivores, natural disasters such as fires and severe winds, and anthropogenic disturbance of soils and plant tissue. At the time of this review, there were no publications characterizing herbivory regimes in the prairie. Anthropogenic disturbance of extant prairies in federally protected patches are associated with the edge conditions generated by current high patch fragmentation. Increasing patch size could help moderate edge conditions enough to favor a broader, less ruderal niche within the patch interior. Further study is required to understand whether ruderal invasives are concentrated at patch edges or if they are more evenly distributed throughout the prairie.

“…it is important to note that contemporary restoration practices such as using prescribed fire may bolster, not hinder, invasibility by exotics.”
In federally unprotected patches, sources of anthropogenic disturbance involve agricultural tilling and land clearing for urban development. Lowering ruderality there would involve the implementation of educational programs to protect unused arable land corridors and minimize unnecessary land clearing during construction.
Finally, it is important to note that contemporary restoration practices such as using prescribed fire may bolster, not hinder, invasibility by exotics. Fire is a source of disturbance. Although fire historically contributed to dispersal filter requirements in some prairie habitats, it appears that other contemporary anthropogenic disturbance sources have increased the overall magnitude of disturbance experienced by these communities, such that providing additional disturbance through prescribed fire would further skew community assembly filters to favor more ruderally proficient species. This is consistent with the results found in one such prescribed fire study (Martin and Hamman, 2016), which recorded a decline in perennial taxa with increasing fire severity.
A recent review (Hamman et al., 2011) of fire use for South Sound prairie restoration suggest that prairies which had historically relied more heavily on anthropogenic disturbance mechanisms for continued persistence would have likely already converted to forest ecosystems during post-glaciation succession were it not for anthropogenic disruption of successional processes. However, the same may not be true for prairies relying more heavily on high stress for persistence. Studies have found that prescribed fires may be detrimental for threatened non-vascular plant species groups within the prairie system (Calabria et al., 2016). For these reasons, it is important to be judicious when deciding to use fire for conservation, and to recognize that mechanisms of ecosystem maintenance do not occur as monolith throughout the region. Conservation approaches should be tailored for each site individually.
Strategy 2: Re-introducing natives with superior ruderal fitness

Clarkia amoena has high metabolic investment in ruderal traits.
As explained in Chapter Four, functional allotment data for species being considered for future introduction could be used to model and predict their functional compatibility with current filter requirements in the community. For example, we would expect that a species candidate with a C-S-R allotment of {80%,10%,10%} would be unlikely to persist in South Sound prairies due to its low compatibility with the current community mean allotment of {22%,37%,40%}. This type of comparative analysis could be used to test the likelihood of success in plant re-introductions before undertaking a major restoration project.
In addition, local functional surveys of plant communities in the vicinity of protected prairie habitats could be conducted to determine the likelihood invasibility success, and to assess invasibility risk per species. This could be used as an invasibility threat assessment tool to focus limited restoration efforts into the most effective areas.
Future Work
Plant survey data on South Sound Prairies are dated and incomplete. A complete plant survey of South Sound Prairies, one that includes both accurate taxonomic richness counts, and relative abundance statistical models for evenness assessment, is long overdue. Investigations on arthropod and mammalian herbivory rate changes could also prove insightful in understanding the prairie’s current disturbance regime.
In addition, there were significant functional data missing in my preliminary analysis. A functional account of Washington’s native flora is urgently needed, such that functional data is available on all endemic species and can be used to inform conservation efforts. Field data collection using leaf traits can be completed with relatively small investment in time and resources using simple tools (Pierce et al., 2013).
References
Altman, B. 2011. Historical and Current Distribution and Populations of Bird Species in Prairie-Oak Habitats in the Pacific Northwest. Northwest Science 85:194–222.
Apostol, D., M. Sinclair, and Society for Ecological Restoration International. 2006. Restoring the Pacific Northwest: the art and science of ecological restoration in Cascadia. Island Press, Washington, DC.
Barnosky, C. W. 1985. Late Quaternary vegetation near Battle Ground Lake, southern Puget Trough, Washington. GSA Bulletin 96:263–271.
Calabria, L. M., K. Petersen, S. T. Hamman, and R. J. Smith. 2016. Prescribed Fire Decreases Lichen and Bryophyte Biomass and Alters Functional Group Composition in Pacific Northwest Prairies. Northwest Science 90:470–483.
Chappell, C. B., M. S. Gee, and B. Stephens. 2003. A geographic information system map of existing grasslands and oak woodlands in the Puget Lowland and Willamette Valley ecoregions, Washington. Washington Natural Heritage Program, Olympia, WA.
Chappell, C., M. Gee, B. Stephens, R. Crawford, and S. Farone. 2001. Distribution and decline of native grasslands and oak woodlands in the Puget Lowland and Willamette Valley ecoregions, Washington. Washington Natural Heritage Program:124–139.
Clements, D. 2013. Translocation of rare plant species to restore Garry oak ecosystems in western Canada: challenges and opportunities. Botany 91.
Crawford, R. C., and H. Hall. 1997. Changes in the South Puget Prairie Landscape. Page 5 Ecology and Conservation of the South Puget Sound Prairie Landscape. The Nature Conservancy.
Diaz, S., J. Kattge, J. H. C. Cornelissen, I. J. Wright, S. Lavorel, S. Dray, B. Reu, M. Kleyer, C. Wirth, I. Colin Prentice, E. Garnier, G. Bönisch, M. Westoby, H. Poorter, P. B. Reich, A. T. Moles, J. Dickie, A. N. Gillison, A. E. Zanne, J. Chave, S. Joseph Wright, S. N. Sheremet’ev, H. Jactel, C. Baraloto, B. Cerabolini, S. Pierce, B. Shipley, D. Kirkup, F. Casanoves, J. S. Joswig, A. Günther, V. Falczuk, N. Rüger, M. D. Mahecha, and L. D. Gorné. 2016. The global spectrum of plant form and function. Nature 529:167–171.
Dorner, J. M. W. 1999. The south Puget Sound prairie plant community: a multivariate analysis of plant species distribution and the relationship of environmental variables. PhD Thesis, University of Washington.
Dunn, P., and K. Ewing. 1997. Ecology and Conservation of the South Puget Sound Prairie Landscape. The Nature Conservancy, Seattle, WA.
Dunwiddie, P. W., E. R. Alverson, R. A. Martin, and R. Gilbert. 2014. Annual Species in Native Prairies of South Puget Sound, Washington. Northwest Science 88:94–105.
Dunwiddie, P. W., E. Alverson, A. Stanley, R. Gilber, S. Pearson, and D. Hays. 2006. The Vascular Plant Flora of the South Puget Sound Prairies, Washington, USA. Davidsonia 14:51–69.
Dunwiddie, P. W., and J. D. Bakker. 2011. The Future of Restoration and Management of Prairie-Oak Ecosystems in the Pacific Northwest. Northwest Science 85:83–92.
Dunwiddie, P. W., and R. A. Martin. 2016. Microsites Matter: Improving the Success of Rare Species Reintroductions. PLoS ONE 11.
Easterly, R., D. Salstrom, and C. B. Chappell. 2005. Wet prairie swales of the South Puget Sound, Washington. The Nature Conservancy, Olympia, WA.
Fazzino, L., H. Elizabeth Kirkpatrick, and C. Fimbel. 2011. Comparison of Hand-Pollinated and Naturally-Pollinated Puget Balsamroot ( Balsamorhiza deltoidea Nutt.) to Determine Pollinator Limitations on South Puget Sound Lowland Prairies. Northwest Science 85:352–360.
Fimbel, C., and S. Freed. 2008. Monitoring Western Gray Squirrels for Landscape Management in Western Washington. Northwest Science 82:299–308.
Floberg, J., M. Goering, G. Wilhere, C. Macdonald, C. Chappell, C. Rumsey, Z. Ferdaña, A. Holt, P. Skidmore, T. Horsman, E. Alverson, C. Tanner, M. Bryer, P. Iachetti, A. Harcombe, B. McDonald, T. Cook, M. Summers, and D. Rolph. 2004. Willamette Valley-PugetTrough-Georgia Basin ecoregional assessment.
Gamon, J., P. W. Dunwiddie, and T. Thomas. 2000. Recovery Plan for the Golden Painbrush. U.S. Fish and Wildlife Service, Portland, OR.
Godt, M. J. W., F. Caplow, and J. L. Hamrick. 2005. Allozyme diversity in the federally threatened golden paintbrush, Castilleja levisecta (Scrophulariaceae). Conservation Genetics 6:87–99.
Haan, N. L., J. D. Bakker, P. W. Dunwiddie, and M. J. Linders. 2018. Instar-specific effects of host plants on survival of endangered butterfly larvae. Ecological Entomology 43:742–753.
Hamman, S., P. Dunwiddie, J. L. Nuckols, and M. McKinley. 2011. Fire as a Restoration Tool in Pacific Northwest Prairies and Oak Woodlands: Challenges, Successes, and Future Directions. Northwest Science 85:317–328.
Li, Y., and B. Shipley. 2017. An experimental test of CSR theory using a globally calibrated ordination method. PLOS ONE 12:e0175404.
Martin, R. A., and S. T. Hamman. 2016. Ignition Patterns Influence Fire Severity and Plant Communities in Pacific Northwest, USA, Prairies. Fire Ecology 12:88–102.
Peter, D. H., and T. B. Harrington. 2014. Historical Colonization of South Puget Sound Prairies by Douglas-Fir at Joint Base Lewis-McChord, Washington. Northwest Science 88:186–205.
Pierce, S., G. Brusa, I. Vagge, and B. E. L. Cerabolini. 2013. Allocating CSR plant functional types: the use of leaf economics and size traits to classify woody and herbaceous vascular plants. Functional Ecology 27:1002–1010.
Pierce, S., D. Negreiros, B. E. L. Cerabolini, J. Kattge, S. Díaz, M. Kleyer, B. Shipley, S. J. Wright, N. A. Soudzilovskaia, V. G. Onipchenko, P. M. van Bodegom, C. Frenette-Dussault, E. Weiher, B. X. Pinho, J. H. C. Cornelissen, J. P. Grime, K. Thompson, R. Hunt, P. J. Wilson, G. Buffa, O. C. Nyakunga, P. B. Reich, M. Caccianiga, F. Mangili, R. M. Ceriani, A. Luzzaro, G. Brusa, A. Siefert, N. P. U. Barbosa, F. S. Chapin III, W. K. Cornwell, J. Fang, G. W. Fernandes, E. Garnier, S. Le Stradic, J. Peñuelas, F. P. L. Melo, A. Slaviero, M. Tabarelli, and D. Tampucci. 2017. A global method for calculating plant CSR ecological strategies applied across biomes world-wide. Functional Ecology 31:444–457.
PLANTS Database | USDA PLANTS. 2020, April 24. . https://plants.sc.egov.usda.gov/java/.
R: The R Project for Statistical Computing. 2020, April 24. . https://www.r-project.org/.
Ren, H., S. Jian, H. Liu, Q. Zhang, and H. Lu. 2014. Advances in the reintroduction of rare and endangered wild plant species. Science China Life Sciences 57:603–609.
Rogers, R., D. Norman, and D. Rolph. 1997. The status of neotropical migrant birds in the prairie landscape. Page 16.
Ryan, L. 1997. Ecology of the Western Gray Squirrel in the South Puget Sound. Page 6 Ecology and Conservation of the South Puget Sound Prairie Landscape. The Nature Conservancy.
Salama, A., M. R. Shukla, E. Popova, N. S. Fisk, M. P. Jones, and P. K. Saxena. 2018. In vitro propagation and reintroduction of golden paintbrush ( Castilleja levisecta), a critically imperilled plant species. Canadian Journal of Plant Science 98:762–770.
Slater, G. L., and B. Altman. 2011. Avian Restoration in the Prairie-Oak Ecosystem: A Reintroduction Case Study of Western Bluebirds to San Juan Island, Washington. Northwest Science 85:223–232.
Stanley, A. G., T. N. Kaye, and P. W. Dunwiddie. 2008. Regional strategies for restoring invaded prairies. Native Plants Journal (Indiana University Press) 9:247–254.
Stinson, D. W. 2005. Washington State status report for the Mazama pocket gopher, streaked horned lark, and Taylor’s checkerspot. Washington Deptof Fish and Wildlife, Olympia, Wash.
Stinson, D. W. 2016. Periodic status review for the Streaked Horned Lark in Washington. Washington Department of Fish and Wildlife, Wildlife Program, Olympia, WA.
Swanson, M. E., N. M. Studevant, J. L. Campbell, and D. C. Donato. 2014. Biological associates of early-seral pre-forest in the Pacific Northwest. Forest Ecology and Management 324:160–171.
Thomas, T., and A. Carey. 1996. Endangered, threatened, and sensitive plants of Fort Lewis, Washington: Distribution, mapping, and management recommendations for species conservation. Northwest Science 70.
US Census Bureau. 2020, March 17. U.S. Population Up 6.3% Since 2010. https://www.census.gov/library/visualizations/interactive/population-increase-2019.html.
Web of Science. 2020, November 11. . Database. https://login.webofknowledge.com/error/Error?Error=IPError&PathInfo=%2F&RouterURL=https%3A%2F%2Fwww.webofknowledge.com%2F&Domain=.webofknowledge.com&Src=IP&Alias=WOK5.
Wright, I. J., P. B. Reich, M. Westoby, D. D. Ackerly, Z. Baruch, F. Bongers, J. Cavender-Bares, T. Chapin, J. H. C. Cornelissen, M. Diemer, J. Flexas, E. Garnier, P. K. Groom, J. Gulias, K. Hikosaka, B. B. Lamont, T. Lee, W. Lee, C. Lusk, and J. J. Midgley. 2004. The worldwide leaf economics spectrum. Nature 428:821–827.